Even though mutualistic interactions are ubiquitous in nature, we are still far from making good predictions about the fate of mutualistic communities under threats such as habitat fragmentation and climate change. Fragmentation often causes declines in abundance of a species due to increased susceptibility to edge effects between remnant habitat patches and lower quality “matrix” surrounding these focal patches. It has been argued that ecological communities are replete with trait-mediated indirect effects, and that these effects may sometimes contribute more to the dynamics of a population than direct density-mediated effects, e.g., lowering an organism’s fitness through competitive interactions. Although some studies have focused on trait-mediated behavior such as trait-mediated dispersal, in which an organism changes its dispersal patterns due to the presence of another species, they have been mostly limited to predator-prey systems–little is known regarding their effect on other interaction systems such as mutualism. Here, we explore consequences of fragmentation and trait-mediated dispersal on coexistence of a system of two mutualists by employing a model built upon the reaction diffusion framework. To distinguish between trait-mediated dispersal and density-mediated effects, we isolate effects of trait-mediated dispersal on the mutualistic system by excluding any direct density-mediated effects in the model. Our results demonstrate that fragmentation and trait-mediated dispersal can have important impacts on coexistence of mutualists. Specifically, one species can be better able to invade and persist than the other and be crucial to the success of the other species in the patch. Matrix quality degradation can also bring about a complete reversal of the role of which species is supporting the other’s persistence in the patch, even as the patch size remains constant. As most mutualistic relationships are identified based on density-mediated effects, such an effect may be easily overlooked.
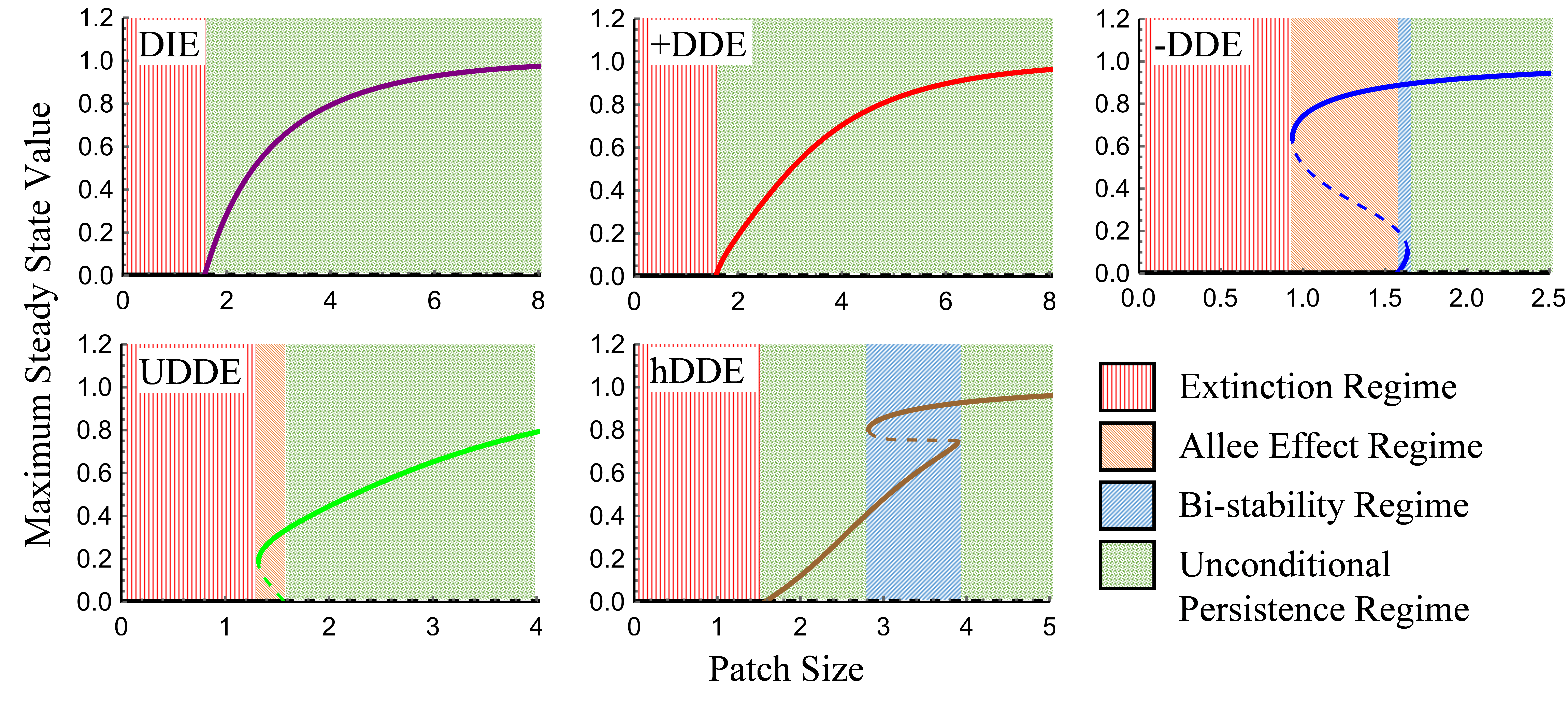
Emigration is a fundamental process affecting species’ local, regional, and large-scale dynamics. The paradigmatic view in ecology is that emigration is density independent (DIE) or positive density dependent (+DDE). However, alternative forms are biologically plausible, including negative (−DDE), U-shaped (uDDE), and hump-shaped (hDDE) forms. We reviewed the empirical literature to assess the frequency of different forms of density-dependent emigration and whether the form depended on methodology. We also developed a reaction-diffusion model to illustrate how different forms of DDE can affect patch-level population persistence. We found 145 studies, the majority representing DIE (30%) and +DDE (36%). However, we also regularly found −DDE (25%) and evidence for nonlinear DDE (9%), including one case of uDDE and two cases of hDDE. Nonlinear DDE detection is likely hindered by the use of few density levels and small density ranges. Based on our models, DIE and +DDE promoted stable and persistent populations. uDDE and −DDE generated an Allee effect that decreases minimum patch size. Last, −DDE and hDDE models yielded bistability that allows the establishment of populations at lower densities. We conclude that the emigration process can be a diverse function of density in nature and that alternative DDE forms can have important consequences for population dynamics.

Covering all major arthropods of medical importance worldwide, this award-winning resource has established itself as a standard reference for almost 25 years. With the globilization of commerce and the world becoming more intimately connected through the everyday ease of travel, unknown arthropod species are being increasingly encountered. This means access to up-to-date, authoritative information in medical entomology has never been more important. Now in its seventh edition, this book maintains its well-acclaimed status as the ultimate easy-to-use guide to identify disease-carrying arthropods, the common signs and symptoms of vector-borne diseases, and the current recommended procedures for treatment.
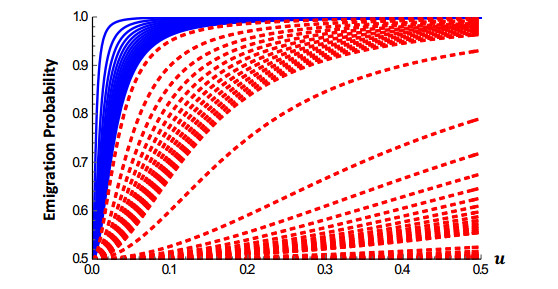

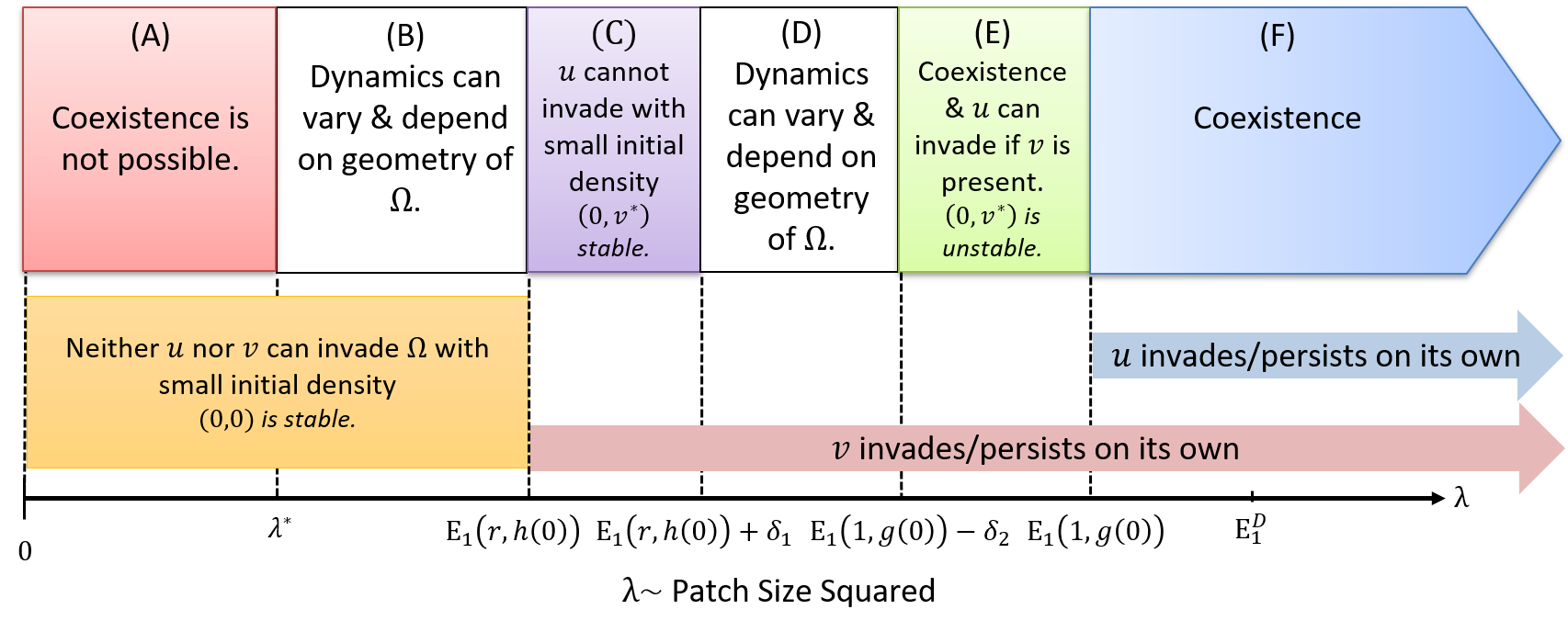
The relationship between conspecific density and the probability of emigrating from a patch can play an essential role in determining the population-dynamic consequences of an Allee effect. In this paper, we model a population that inside a patch is diffusing and growing according to a weak Allee effect per-capita growth rate, but the emigration probability is dependent on conspecific density. The habitat patch is one-dimensional and is surrounded by a tuneable hostile matrix. We consider five different forms of density dependent emigration (DDE) that have been noted in previous empirical studies. Our models predict that at the patch-level, DDE forms that have a positive slope will counteract Allee effects, whereas, DDE forms with a negative slope will enhance them. Also, DDE can have profound effects on the dynamics of a population, including producing very complicated population dynamics with multiple steady states whose density profile can be either symmetric or asymmetric about the center of the patch. Our results are obtained mathematically through the method of subsuper solutions, time map analysis, and numerical computations using Wolfram Mathematica.
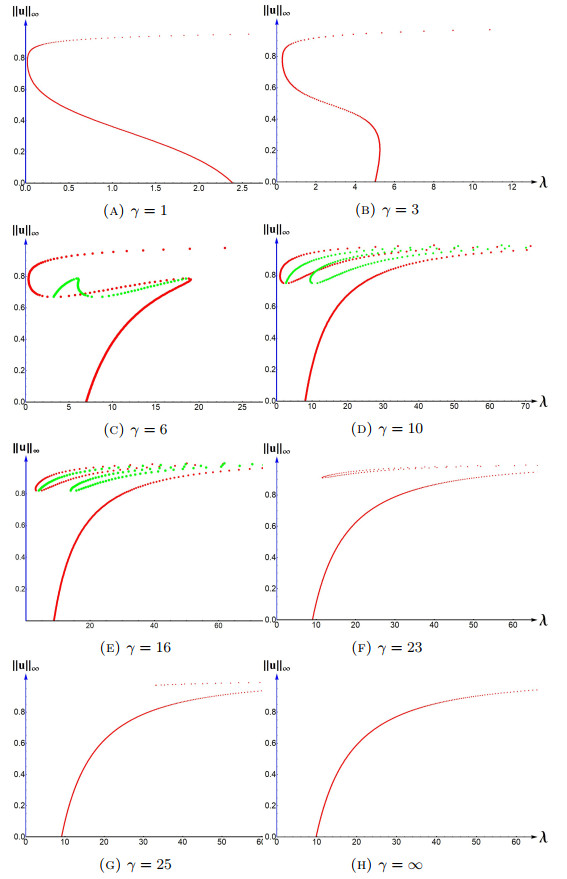
We study positive solutions to a steady state reaction diffusion equation arising in population dynamics, namely, \begin{equation*} \label{abs} \left\lbrace \begin{matrix}-\Delta u=\lambda u(1-u) ;~x\in\Omega\\ \frac{\partial u}{\partial \eta}+\gamma\sqrt{\lambda}[(A-u)^2+\epsilon]u=0; ~x\in\partial \Omega \end{matrix} \right. \end{equation*} where \(\Omega\) is a bounded domain in \(\mathbb{R}^N\); \(N > 1\) with smooth boundary \(\partial \Omega\) or \(\Omega=(0,1)\), \(\frac{\partial u}{\partial \eta}\) is the outward normal derivative of \(u\) on \(\partial \Omega\), \(\lambda\) is a domain scaling parameter, \(\gamma\) is a measure of the exterior matrix (\(\Omega^c\)) hostility, and \(A\in (0,1)\) and \(\epsilon > 0\) are constants. The boundary condition here represents a case when the dispersal at the boundary is U-shaped. In particular, the dispersal is decreasing for \(u < A\) and increasing for \(u > A\). We will establish non-existence, existence, multiplicity and uniqueness results. In particular, we will discuss the occurrence of an Allee effect for certain range of \(\lambda\). When \(\Omega=(0,1)\) we will provide more detailed bifurcation diagrams for positive solutions and their evolution as the hostility parameter \(\gamma\) varies. Our results indicate that when \(\gamma\) is large there is no Allee effect for any \(\lambda\). We employ a method of sub-supersolutions to obtain existence and multiplicity results when \(N > 1\), and the quadrature method to study the case \(N=1\).
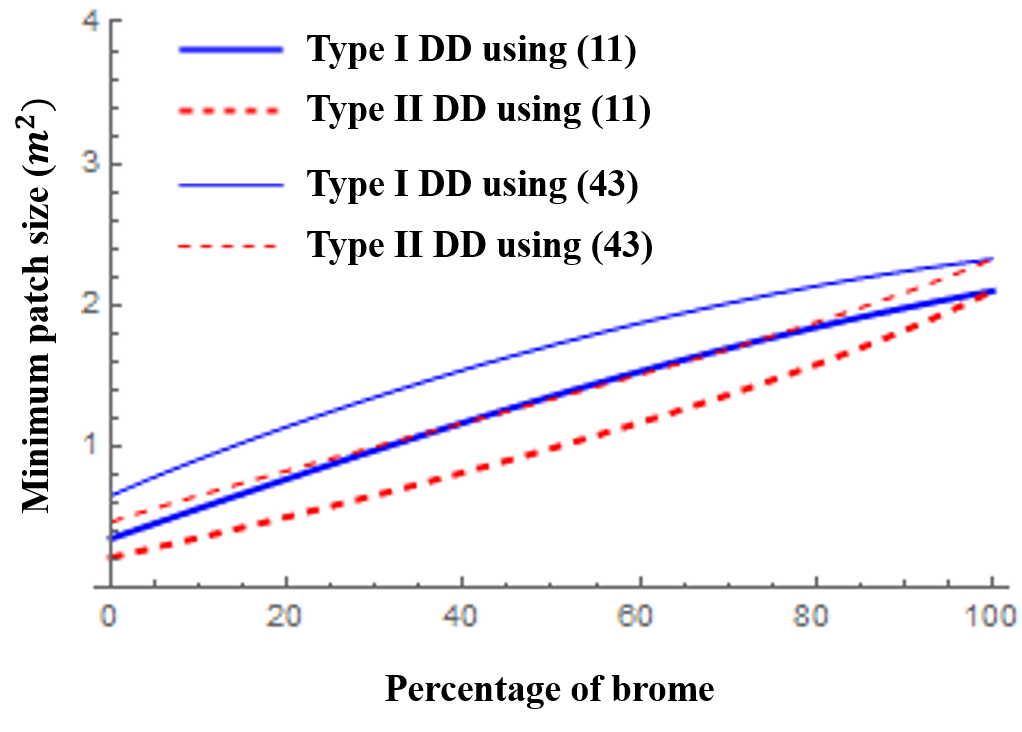
Fragmentation creates landscape-level spatial heterogeneity which in turn influences population dynamics of the resident species. This often leads to declines in abundance of the species due to increased susceptibility to edge effects between the remnant habitat patches and the lower quality “matrix” surrounding these focal patches. In this paper, we formalize a framework to facilitate the connection between small-scale movement and patch-level predictions of persistence through a mechanistic model based on reaction–diffusion equations. The model is capable of incorporating essential information about edge-mediated effects such as patch preference, movement behavior, and matrix-induced mortality. We mathematically analyze the model’s predictions of persistence with a general logistic-type growth term and explore their sensitivity to demographic attributes in both the patch and matrix, as well as patch size and geometry. Also, we provide bounds on demographic attributes and patch size in order for the model to predict persistence of a species in a given patch based on assumptions on the patch/matrix interface. Finally, we illustrate the utility of this framework with a well-studied planthopper species (Prokelisia crocea) living in a highly fragmented landscape. Using experimentally derived data from various sources to parameterize the model, we show that, qualitatively, the model results are in accord with experimental predictions regarding minimum patch size of P. crocea. Through application of a sensitivity analysis to the model, we also suggest a ranking of the most important model parameters based on which parameter will cause the largest output variance.
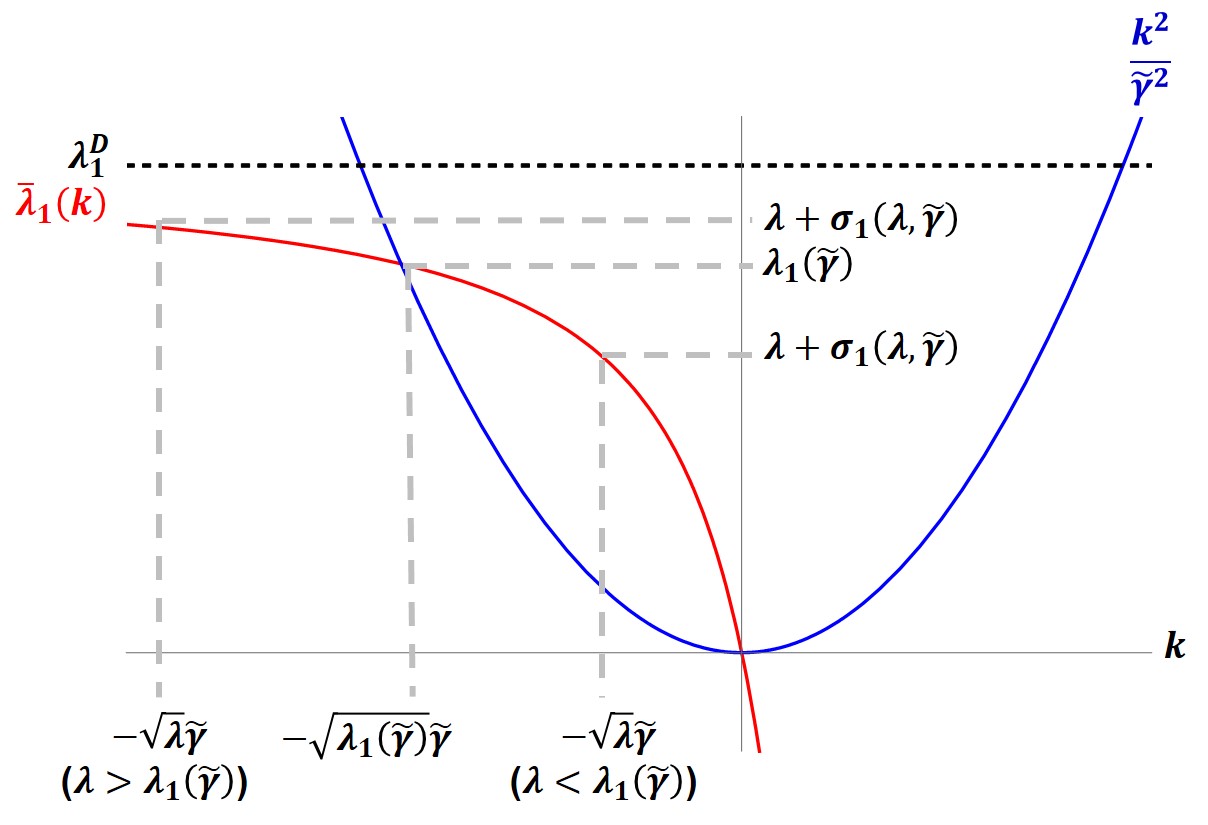
We study positive solutions to the steady state reaction diffusion equation: \begin{equation*} \begin{cases} - \Delta v = \lambda v(1-v), & x \in \Omega_0, \\ \frac{\partial v}{\partial \eta} + \gamma \sqrt{\lambda} ( v-A)^2 v =0 , & x \in \partial \Omega_0, \end{cases} \end{equation*} where $\Omega_0$ is a bounded domain in $\mathbb{R}^n$; $n \ge 1$ with smooth boundary $\partial \Omega_0$, ${\partial }/{\partial \eta}$ is the outward normal derivative, $A \in (0,1)$ is a constant, and $\lambda$, $\gamma$ are positive parameters. Such models arise in the study of population dynamics when the population exhibits a U-shaped density dependent dispersal on the boundary of the habitat. We establish existence, multiplicity, and uniqueness results for certain ranges of the parameters $\lambda$ and $\gamma$. We obtain our existence and mulitplicity results via the method of sub-super solutions.
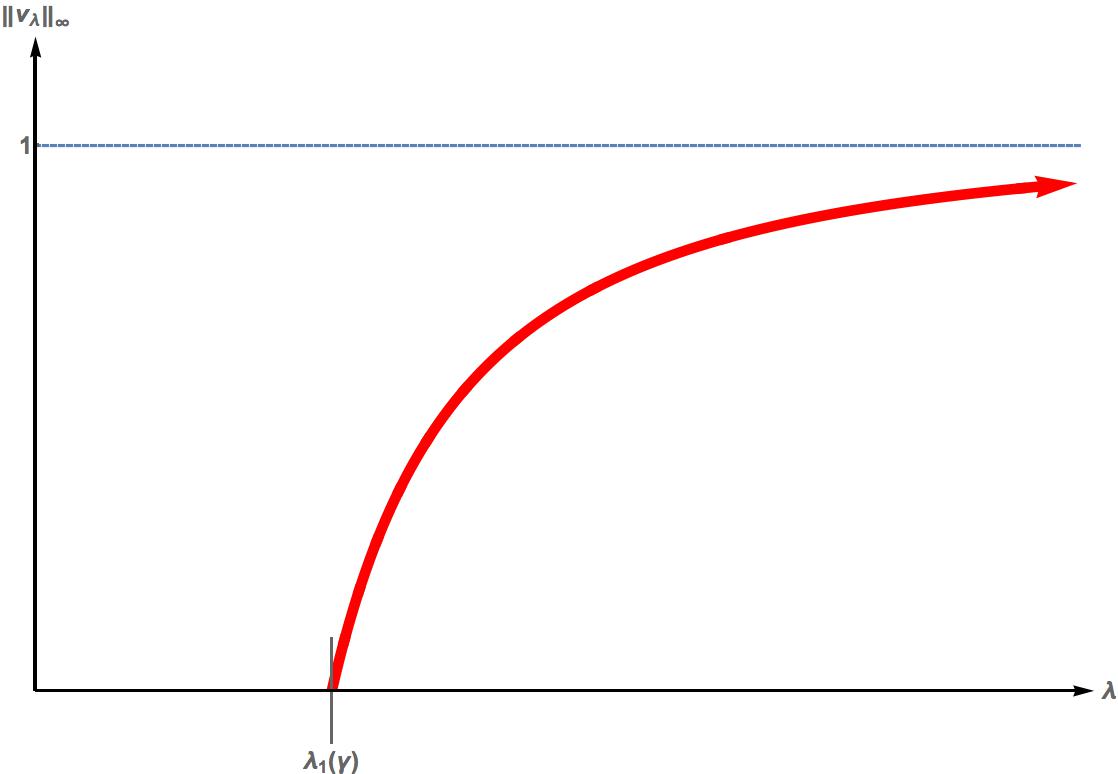
We analyze the positive solutions to \begin{equation*} \left\{ \begin{array}{cl} - \Delta v = \lambda v(1-v); & \Omega_0, \\ \frac{\partial v}{\partial \eta} + \gamma \sqrt{\lambda} v =0 ; & \partial \Omega_0, \end{array} \right. \end{equation*} where $\Omega_0=(0,1)$ or is a bounded domain in $\mathbb{R}^n$; $n =2,3$ with smooth boundary and $|\Omega_0|=1$, and $\lambda, \gamma$ are positive parameters. Such steady state equations arise in population dynamics encapsulating assumptions regarding the patch/matrix interfaces such as patch preference and movement behavior. In this paper, we will discuss the exact bifurcation diagram and stability properties for such a steady state model.

We discuss a quadrature method for generating bifurcation curves of positive solutions to some autonomous boundary value problems with nonlinear boundary conditions. We consider various nonlinearities, including positone and semipositone problems in both singular and nonsingular cases. After analyzing the method in these cases, we provide an algorithm for the numerical generation of bifurcation curves and show its application to selected problems.
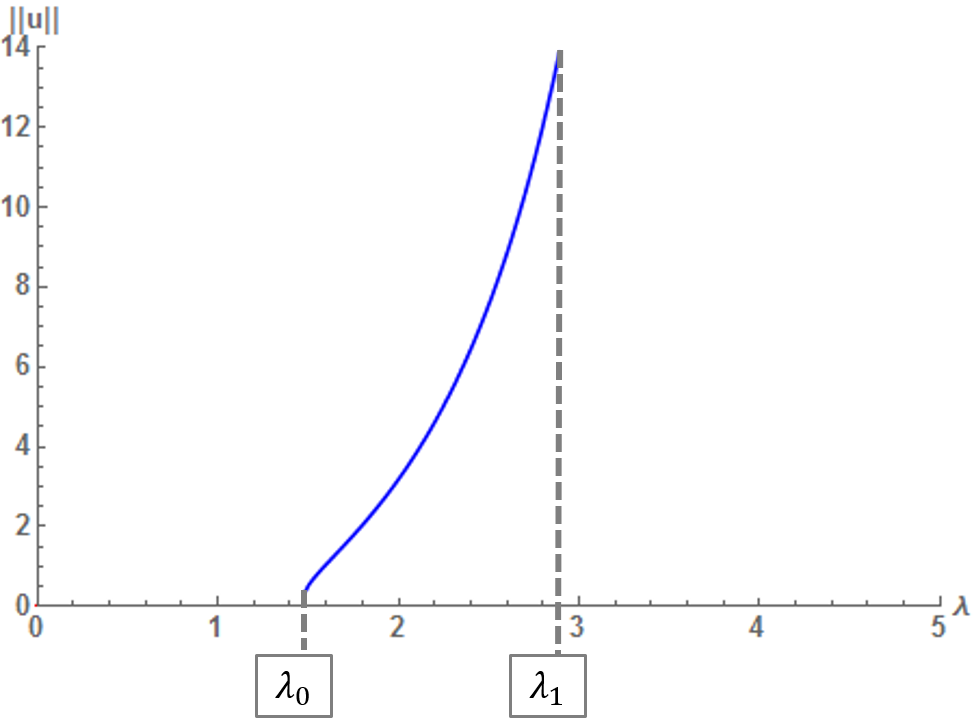
We investigate the stability properties of positive steady-state solutions of semilinear initial–boundary-value problems with nonlinear boundary conditions. In particular, we employ a principle of linearized stability for this class of problems to prove sufficient conditions for the stability and instability of such solutions. These results shed some light on the combined effects of the reaction term and the boundary nonlinearity on stability properties. We also discuss various examples satisfying our hypotheses for stability results in dimension 1. In particular, we provide complete bifurcation curves for positive solutions for these examples.

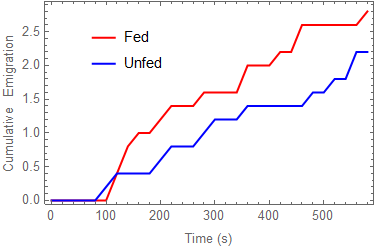
Bed bug problems have been increasing since the 1980s, and accordingly, there have been intensive efforts to better understand their biology and behavior for control purposes. Understanding bed bug diffusion rates and dispersal patterns from one site to another (or lack thereof) is a key component in prevention and control campaigns. This study analyzed diffusion rates and dispersal patterns in a population of bed bugs, recently fed and unfed, in both one-dimensional and two-dimensional settings. When placed in the middle of a 71 cm × 2.7 cm artificial lane, approximately half of the bugs regardless of feeding status stayed at or near the release point during the 10 min observation periods, while about a fourth of them walked to the end of the lane. When placed in the middle of an arena measuring 51 cm × 76 cm and allowed to walk in any direction, approximately one-fourth of bed bugs, fed or unfed, still remained near their release point (no significant difference between fed or unfed). As for long-distance dispersal, 11/50 (22%) of recently fed bed bugs moved as far as possible in the arena during the 10 min replications, while only 2/50 (4%) unfed bed bugs moved to the maximum distance. This difference was significantly different (p < 0.0038), and indicates that unfed bed bugs did not move as far as recently fed ones. A mathematical diffusion model was used to quantify bed bug movements and an estimated diffusion rate range of 0.00006 cm2/s to 0.416 cm2/s was determined, which is almost no movement to a predicted root mean squared distance of approximately 19 cm per 10 min. The results of this study suggest that bed bugs, upon initial introduction into a new area, would have a difficult time traversing long distances when left alone to randomly disperse.
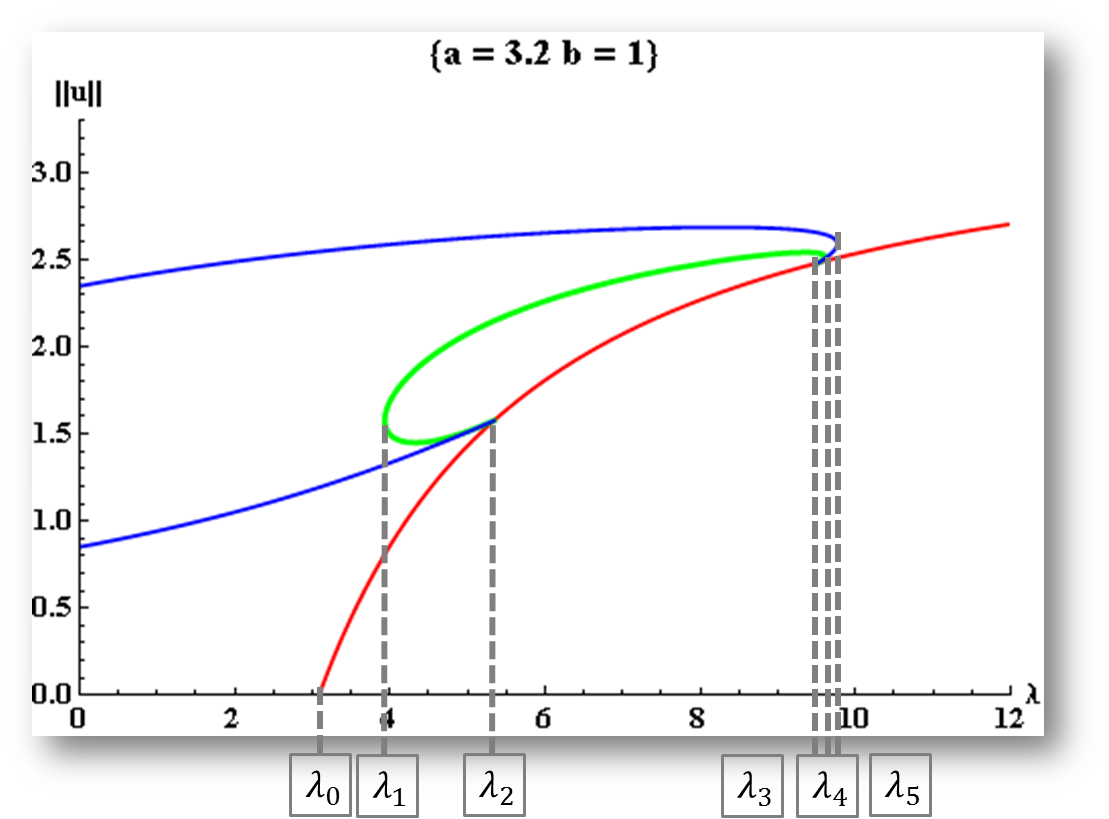

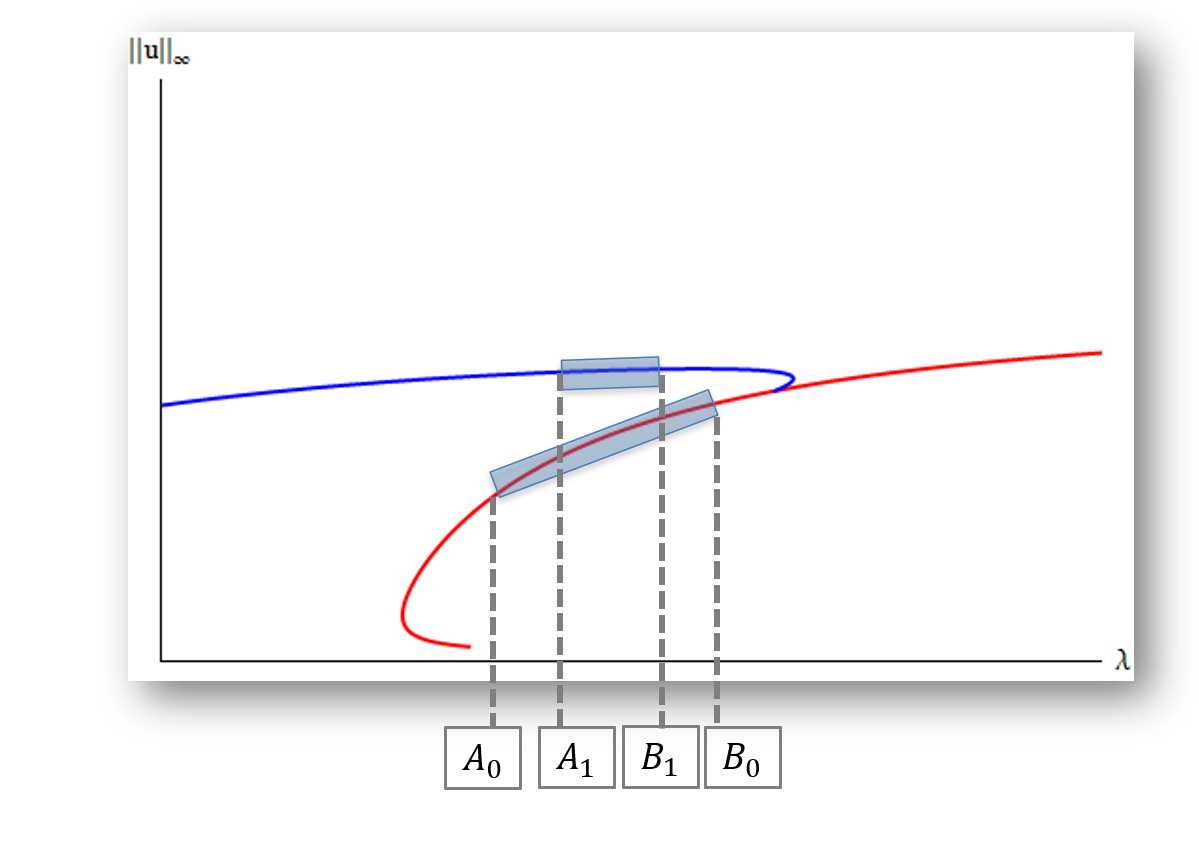
We examine the structure of positive steady state solutions for a diffusive population model with logistic growth and negative density dependent emigration on the boundary. In particular, this class of nonlinear boundary conditions depends on both the population density and the diffusion coefficient. Results in the one-dimensional case are established via quadrature methods. Additionally, we discuss the existence of a Halo-shaped bifurcation curve.
The structure of positive steady state solutions of a diffusive logistic population model with constant yield harvesting and negative density dependent emigration on the boundary is examined. In particular, a class of nonlinear boundary conditions that depends both on the population density and the diffusion coefficient is used to model the effects of negative density dependent emigration on the boundary. Our existence results are established via the well-known sub-super solution method.
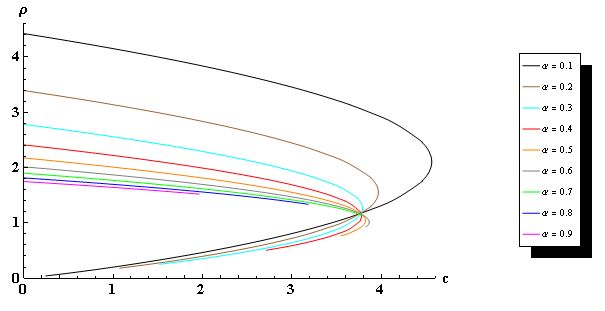
We consider the problem \begin{equation*} \label{eqn} \left\{ \begin{split} -\Delta u &=\frac{au-bu^2-c}{u^\alpha} , \quad x \in \Omega \\u &= 0, \qquad \qquad \qquad \ x \in \partial\Omega \end{split} \right. \end{equation*} where $\Omega$ is a smooth bounded domain in $\mathbb{R}^{n}$, $a>0, b>0, c\geq0$ and $\alpha \in $ $(0, 1)$. Given $a,b$ and $\alpha,$ we establish the existence of a positive solution for small values of $c$. We also extend our results to the $\Delta_p$ operator and to corresponding exterior domain problems.

We examine a one-dimensional reaction diffusion model with a weak Allee growth rate that appears in population dynamics. We combine grazing with a certain nonlinear boundary condition that models negative density dependent dispersal on the boundary and analyze the effects on the steady states. In particular, we study the bifurcation curve of positive steady states as the grazing parameter is varied. Our results are acquired through the adaptation of a quadrature method and Mathematica computations. Specifically, we computationally ascertain the existence of Σ-shaped bifurcation curves with several positive steady states for a certain range of the grazing parameter.

We analyze the solutions of a population model with diffusion and strong Allee effect. In particular, we focus our study on a population that satisfies a certain nonlinear boundary condition and on its survival when constant yield harvesting is introduced. We discuss, in detail, results for the one-dimensional case.

We consider a population model with diffusion, a strong Allee effect per capita growth function, and constant yield harvesting. In particular, we focus our study on a population living in a patch, $\Omega \subseteq \mathbb{R}^n$ with $n \geq 1$, that satisfies a certain nonlinear boundary condition. We establish our existence results by the method of sub-super solutions.
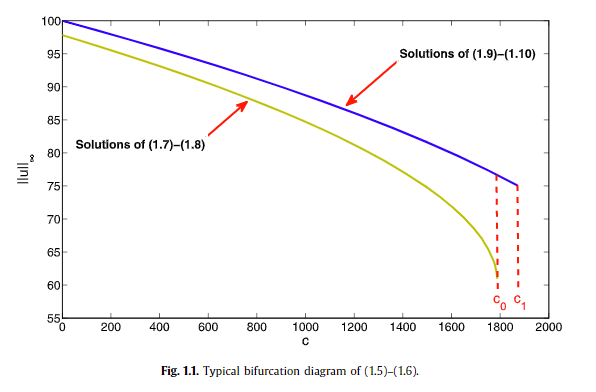
We analyze the solutions of a population model with diffusion and logistic growth. In particular, we focus our study on a population living in a patch, $\Omega \subset \mathbb{R}^n$ with $n \geq 1$, that satisfies a certain non-linear boundary condition and on its survival when constant yield harvesting is introduced. We establish our existence results by the method of sub-super solutions.
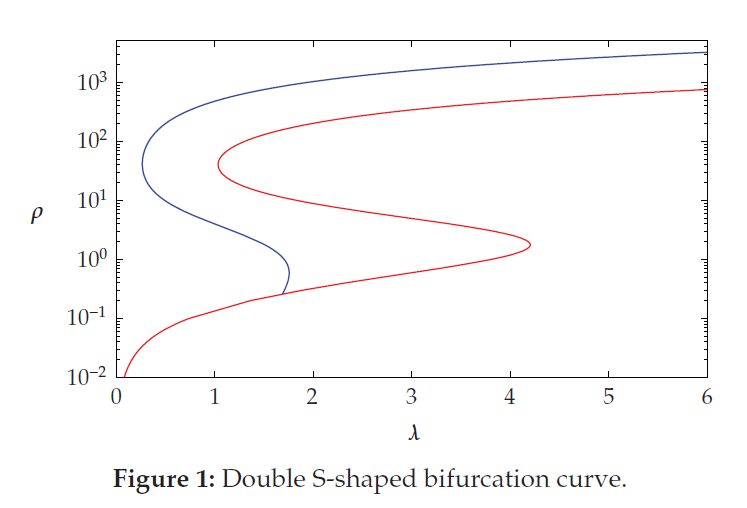
We study the positive solutions to boundary value problems of the form \begin{eqnarray*} -\Delta u & = & \lambda f(u); \quad \Omega\\ \alpha(x, u)\frac{\partial u}{\partial \eta} & + & \left[1 - \alpha(x, u) \right]u = 0; \quad \partial \Omega \end{eqnarray*} where $\Omega$ is a bounded domain in $\mathbb{R}^n$ with $n \geq 1$, $\Delta$ is the Laplace operator, $\lambda$ is a positive parameter, $f:[0, \infty) \longrightarrow (0, \infty)$ is a continuous function which is sublinear at $\infty$, $\frac{\partial u}{\partial \eta}$ is the outward normal derivative, and $\alpha(x, u):\Omega \times \mathbb{R} \longrightarrow [0, 1]$ is a smooth function nondecreasing in $u$. In particular, we discuss the existence of at least two positive radial solutions for $\lambda \gg 1$ when $\Omega$ is an annulus in $\mathbb{R}^n.$ Further, we discuss the existence of a double S-shaped bifurcation curve when $n = 1$, $\Omega = (0, 1)$, and $f(s) = e^{\frac{\beta s}{\beta + s}}$ with $\beta \gg 1.$

We study a two point boundary-value problem describing the steady states of a Logistic growth population model with diffusion and constant yield harvesting. In particular, we focus on a model when a certain nonlinear boundary condition is satisfied.


Whether or not people get tick-borne diseases (TBD) is related to tick population numbers (the vector), tick infection rates (the pathogen), and exposure of humans and other animals (the hosts). Understanding each of these components and their interplay is important in choosing personal protection measures against ticks as well as prevention/management of TBD. A way of estimating how many ticks occur in a given area would be of great interest to public health officials, as well as how many might actually get on a person during outdoor activities. Tick populations might be sampled and counted directly in small areas, or estimated in larger expanses by using mark-release-recapture (MRR) methods (Daniels et al. 2000, Goddard and Goddard 2008). Collection methods for these MRR studies have included dragging a white cloth around in the woods (drag cloth sampling) and/or walk-around surveys. By using these data, researchers have attempted to design ecological or entomologic risk maps or charts for Lyme disease and other TBD (Davis et al. 1984, Schulze et al. 1991, Daniels et al. 1998, Supergan and Karbowiak 2009). This pilot study attempts to link drag-cloth sampling results to the actual risk of acquiring black-legged ticks, Ixodes scapularis in central Mississippi.
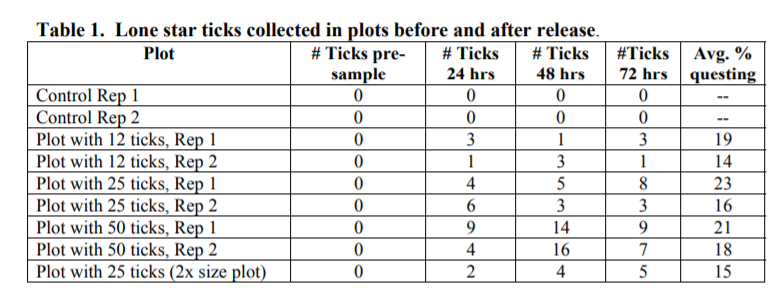
To assess the proportion of lone star ticks questing in an area, a predetermined number of ticks were released into each of 9 wooded/grassy plots in central Mississippi that were known to be free of lone star ticks. Plots were 3 x 15 m and contained 0 (control), 12, 25, and 50 ticks, with two replicates of each. The ninth plot was 2x the size of the others and contained 25 ticks. Plots were then sampled at 3:00 p.m. each day with a 1m2 drag cloth at 24, 48, and 72 hour intervals after tick release. Out of the 199 ticks released, 29 (14.5%) were recaptured at 24 hours, 46 (23.1%) were recaptured at 48 hours, and 36 (18.0%) were recaptured at 72 hours. Size of the plot made little difference in number of ticks collected; the one double-sized plot containing 25 ticks produced sampling results similar to the other, standardsized plots containing the same number of ticks. There were no significant differences in percent tick questing (PTQ) between sites (plots) or times. There was also little relationship between temperature and humidity and PTQ in this study. A logistic regression on the data set showed no significant relationship between tick populations and fraction of ticks questing.
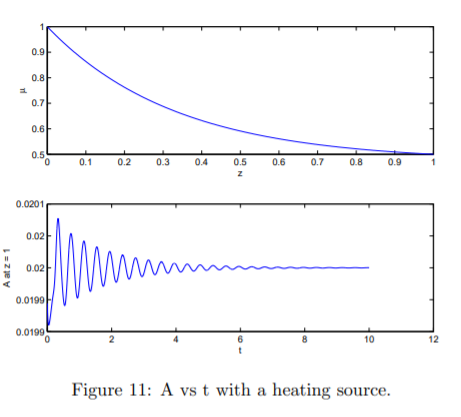
The goal of this research is to revisit the stability results of Yeow (1974) on extending flows with free surfaces. The eigenvalue problem is formulated and solved for the flow of a Newtonian film such as the one encountered in film casting. The stable and unstable region boundaries is obtained. The analysis is extended to a non-isothermal case similar to the work by Shah and Pearson (1972) and German et al. (2006). Stability boundaries for different attenuation velocities (atthe inlet and the exit of the process), and viscosity ratios are obtained. For the solutions which are unstable (or marginally unstable), timedependent solutions (oscillating or growing in time) describing the free surface motion are obtained. The analysis which is applicable to a filament geometry is extended to a sheet with two length scales.

A sequential Bayesian algorithm and accompanying computer program were developed and validated to estimate population numbers of adult blacklegged tick, Ixodes scapularis Say, using mark-release-recapture methodology in field plots in central Mississippi. In fieldwork, data taken in February 2005 in a 1-ha plot yielded an estimate of 317 adult I. scapularis per ha Data from another field plot in 2006, 3 km away, yielded an estimate of 280 adult I. scapularis per ha The number of ticks collected per hour in both plots averaged 4.5. In eight of 14 (57%) of sampling events, the number of ticks collected per hour hovered closely around 5.0 (4.8-5.3). The computer program developed in this study readily produced statistical measures such as mean number of ticks per plot, mode, variance, and standard deviation, as well as easy-to-read graphs of estimated tick populations for each sampling period.

